In summers 2018 and 2019, Amy mapped and collected leaf tissue from all individuals in the study areas and harvested heads from a subset of Echinacea individuals at populations in the NW corner of the Echinacea Project study area (populations: ALF, EELR, KJ, NWLF, GC, SGC, NGC, KJ, NNWLF) to map pollen movement (see Reproductive Fitness in Remnants). Amy analyzed patterns of gene flow, by assessing how individuals’ location and timing of flowering influence their reproductive success and distance of pollen movement. The now four-year-old seedlings are planted in p10.
There was no fieldwork conducted this summer but, in exciting news, Amy used the seed set data from the gene flow in remnants experiment in her dissertation chapter, “Variation in reproductive fitness among individual plants depends on the spatial proximity of prospective mates and the timing of their reproduction”. Amy defended her dissertation in May, and this chapter is currently in preparation for publication!
Amy visited the station a few times this summer! Cupcakes in the field to celebrate a successful dissertation defense!
Start year: 2018
Location: Roadsides, railroad rights of way, and nature preserves in and around Solem Township, MN
In summers 2018 and 2019, I mapped and collected leaf tissue from all individuals in the study areas and harvested seedheads from a subset of Echinacea individuals at populations in the NW corner of the study area (populations: ALF, EELR, KJ, NWLF, GC, SGC, NGC, KJ, NNWLF) to map pollen movement (see Reproductive Fitness in Remnants). To analyze patterns of gene flow, I will assess how individuals’ location and timing of flowering influence their reproductive success and distance of pollen movement. I am currently wrapping up genotyping the DNA from the leaf tissue samples and a subset of the seeds I collected. This summer, the team measured the 3-year-old seedlings from the gene flow study that are planted in exPt10. I did not do additional field work for this project this year.
A flowering Echinacea angustifolia
Start year: 2018
Location: Roadsides, railroad rights of way, and nature preserves in and around Solem Township, MN
Data collected: exPt10 measure data is in the cgdata repo.
Products: I presented a poster based on the locations and flowering phenology of individuals from summer 2018 at the International Pollinator Conference in Davis, CA this summer. The poster is linked here: https://echinaceaproject.org/international-pollinator-conference/.
On Friday, grad student Amy W. paid a visit to the lab at the Chicago Botanic Garden to x-ray Echinacea achenes for several of her projects including the Dust Project, interremnant crosses, and gene flow experiments. We’re thrilled to have a functional x-ray machine once again. Amy noticed lots of variation in her samples, so we’re excited to learn about seed set for these experiments!
In summer 2021, Amy Waananen continued the interremnant crosses experiment to understand how the distance between plants in space and in their timing of flowering influences the fitness of their offspring. This experiment builds on Amy’s study of gene flow and pollen movement in the remnants, asking the question of how pollen movement patterns affect offspring establishment and fitness. If plants that are located close together or flower at the same time are closely related, their offspring might be more closely related and inbred, and have lower fitness than plants that are far apart and/or flower more asynchronously. In other words, if distance in space or time is correlated with relatedness, we’d expect mating between more distant or asynchronous individuals to result in more fit offspring.
To test this hypothesis, Amy performed crosses between plants across a range of spatial isolation (within the same population, in adjacent populations, and in far-apart populations) in 2020. With the team’s help, she also kept track of the individuals’ flowering time to assess whether reproductive synchrony is associated with reduced offspring fitness, suggesting that individuals that flower at the same time are more closely related.
In 2021, Amy repeated the same hand crossing methods to assess the fitness consequences of outcrossing, this year on 44 focal plants.
To learn more about Amy’s project, check out this video created by RET participant Alex Wicker.
Amy collects pollen from Echinacea anthers
Start year: 2020
Location: On27, SGC, GC, NGC, EELR, KJ, NNWLF, NWLF, LF
Data/Materials collected: Style shriveling and seed set and weight from crosses, start and end date of flowering, coordinates of all individuals in the populations listed above
During the summer of 2019, Team Echinacea planted over 1400 E. angustifolia seedlings into 12 plots in a prairie restoration at West Central Area High School in Barrett, MN. We planted seedlings from three sources: (1) offspring from exPt1, (2) plants from my gene flow experiment, and (3) offspring from the Big Event. In summer 2021, Drake also planted plugs of other species (pictured below).
This summer, the team measured the 2-year old seedlings from my gene flow study in exPt10, as well as a few seedlings from the other plantings within the plot. The seedlings from my gene flow experiment are the offspring of open-pollinated Echinacea in 9 populations in the study area. I am assessing the paternity of these seedlings to understand contemporary pollen movement patterns within and among the remnants. In summer 2018, I mapped and collected leaf tissue from all Echinacea individuals within 800m of the study areas and harvested seedheads from a sample of these individuals (see Reproductive Fitness in Remnants). In spring 2019, I germinated and grew up a sample of the seeds that I harvested to obtain leaf tissue for genotyping.
Then, with the team’s help, I planted these seedlings in exPt10 in June 2019. I also collected seeds and leaf tissue in summer 2019 to repeat this process, but I did not germinate the achenes in the following spring because I was not able to assess seed set due to the broken x-ray machine at the CBG and then COVID-related restrictions. I hope to germinate those this spring and plant in summer 2022. I am working on extracting the DNA from the leaf tissue samples I have, which I will use to match up the genotypes of the offspring (i.e., the seeds) with their most likely father (i.e., the pollen source).
A sampler platter of seedlings, planted as part of Drake’s study of how prairie communities respond to parasitic plants.
Start year: 2018
Location: West Central Area High School’s Environmental Learning Center, Barrett, MN, Remnant prairies in Solem Township, Minnesota
This summer I started a new experiment to understand how the distance between plants in space and in their timing of flowering influences the fitness of their offspring. This experiment builds on my study of gene flow and pollen movement in the remnants, asking the question of how pollen movement patterns affect offspring establishment and fitness. If plants that are located close together or flower at the same time are closely related, their offspring might be more closely related and inbred, and have lower fitness than plants that are far apart and/or flower more asynchronously. In other words, if distance in space or time is correlated with relatedness, we’d expect mating between more distant or asynchronous individuals to result in more fit offspring.
To test this hypothesis, I performed crosses between plants across a range of spatial isolation (within the same population, in adjacent populations, and in far-apart populations). With the team’s help, I also kept track of the individuals flowering time so that I can assess whether reproductive synchrony is associated with reduced offspring fitness, suggesting that individuals that flower at the same time are more closely related.
I ended up using 42 focal plants (two of which were mowed before I could harvest them) and a total of 167 sires. I planted 359 offspring from these crosses in November. Next spring and summer, I will measure the seedlings to collect data on emergence and growth. Seed set was lower than I wanted it to be (only ~20%, when I would have expected 60-70% based on compatibility rates in the remnants), so I will also likely perform more crosses in summer 2021 to shore up my sample size.
Crossing at scenic On 27
Start year: 2020
Location: On27, SGC, GC, NGC, EELR, KJ, NNWLF, NWLF, LF
Data/Materials collected: 40 seedheads, style shriveling and seed set and weight from crosses, start and end date of flowering, coordinates of all individuals in the populations listed above
Products: I planted the seeds from the crosses in a plot adjacent to P1 in November, as detailed in this flog post.
During the summer of 2019, Team Echinacea planted over 1400 E. angustifolia seedlings into 12 plots in a prairie restoration at West Central Area High School in Barrett, MN. We planted seedlings from three sources: (1) offspring from exPt1, (2) plants from my gene flow experiment, and (3) offspring from the Big Event. To test how different fire regimes affect fitness in Echinacea, folks from West Central Area plan to apply regular fall burn treatments to four plots, regular spring burn treatments to four other plots, and the remaining four plots will not be burned. I’m not sure if they were able to perform these burns as planned in Fall 2020 given COVID restrictions this spring and fall, but John Van Kempen would be the man to ask about that. I believe they were able to do the burns in the spring.
This summer, the team measured the 1-year old seedlings from my gene flow study in exPt10, as well as a few seedlings from the other plantings within the plot. The seedlings from my gene flow experiment are the offspring of open-pollinated Echinacea in 9 populations in the study area. I am assessing the paternity of these seedlings to understand contemporary pollen movement patterns within and among the remnants. In summer 2018, I mapped and collected leaf tissue from all Echinacea individuals within 800m of the study areas and harvested seedheads from a sample of these individuals (see Reproductive Fitness in Remnants). In spring 2019, I germinated and grew up a sample of the seeds that I harvested to obtain leaf tissue for genotyping.
Then, with the team’s help, I planted these seedlings in exPt10 in June 2019. I also collected seeds and leaf tissue in summer 2019 to repeat this process, but I did not germinate the achenes in the following spring because I was not able to assess seed set due to the broken x-ray machine at the CBG and then COVID-related restrictions. I hope to germinate those this spring and plant in summer 2021. I am working on extracting the DNA from the leaf tissue samples I have, which I will use to match up the genotypes of the offspring (i.e., the seeds) with their most likely father (i.e., the pollen source).
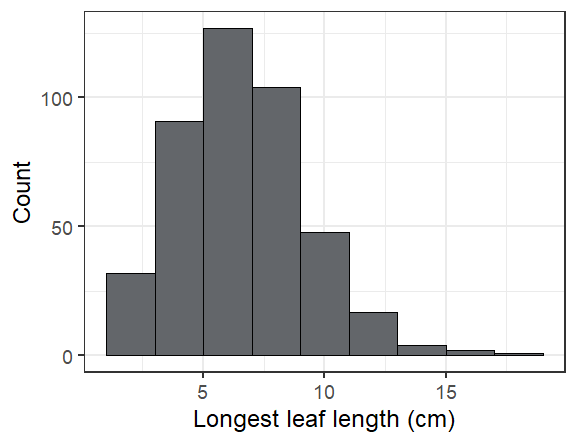
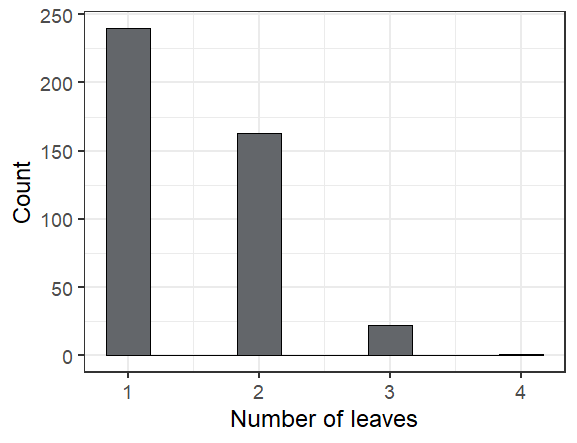
Here are some fun facts about the seedlings we found in exPt 10:
The longest leaf we saw was 19 cm! Most were much smaller (see below).
The leafiest plant we saw had 4 leaves (though one had been munched)
Overall we found 424 seedlings alive of the 598 that we searched for, or 71%. The ones we didn’t find are probably dead, but we’ll look for them again next year to make sure we didn’t just miss them.
I’m looking forward to seeing these friends again next year.
Allie gives a thumbs after successfully finding a baby Echinacea plant in p10!
Start year: 2018
Location: West Central Area High School’s Environmental Learning Center, Barrett, MN, Remnant prairies in Solem Township, Minnesota
In summer
2019, I completed a second season of field work for a study monitoring pollen
movement between remnant populations. In summer 2018, I chose two focal areas,
the NW sites in the study area (populations: ALF, EELR, KJ, NWLF, GC, SGC, NGC,
KJ, NNWLF) and SW sites (populations: LC, NRRX, RRX, YOH, and two large populations
in between these sites). This summer, I limited the study to the NW sites. As
in 2018, I mapped and collected leaf tissue from all individuals in the study
areas and harvested seedheads from a subset of these individuals (see Reproductive Fitness in Remnants). In addition, I monitored the
flowering phenology of all of the flowering plants in these populations (see
Phenology in the Remnants).
Now, I am
working on extracting and genotyping the DNA from the leaf tissue samples and a
subset of the seeds I collected. This takes a long time! I will use the
microsatellite markers that Jennifer Ison developed in her dissertation to
match up the genotypes of the offspring (i.e., the seeds) with their most
likely father (i.e., the pollen source). To analyze patterns of gene flow, I
will assess how individuals’ location and timing of flowering influence their
reproductive success and distance of pollen movement.
In
addition, last summer we planted all of the seedlings from 2018 in the
experimental plot that John Van Kempen set up at West Central Area High School.
We will continue to monitor these seedlings to understand how pollen movement
distance (or the distance between parents) influences offspring fitness.
Here is the team after we planted nearly 298 seedlings in the experimental plot at WCA!
Start
year: 2018
Location: Roadsides, railroad rights of way, and nature preserves in and around
Solem Township, MN
Products: I presented a poster based on
the locations and flowering phenology of individuals from summer 2018 at the
International Pollinator Conference in Davis, CA this summer. The poster is
linked here: https://echinaceaproject.org/international-pollinator-conference/.
In summer 2018, I began a project to look at pollen movement within and among the remnant populations. To do this, I chose two focal areas, the NW sites in the study area (populations: ALF, EELR, KJ, NWLF, GC, SGC, NGC, KJ, NNWLF) and the SW sites (populations: LC, NRRX, RRX, YOH, and two large populations in between these sites). I mapped and collected leaf tissue from all individuals in the study areas and harvested seedheads from a subset of these individuals (see Reproductive Fitness in Remnants). I am currently extracting the DNA from the leaf tissue samples and a subset of the seeds I collected, and will use the microsatellite markers that Jennifer Ison developed in her dissertation to match up the genotypes of the offspring (i.e., the seeds) with their most likely father (i.e., the pollen source).
An Echinacea that has had today’s load of pollen fully removed by pollinators