A rainbow-like cloud glows over exPt01 while the team was in MN for fall 2023 burns. A good omen for the field season that followed!
Introduction
Every year since 1996, members of Team Echinacea have recorded flowering phenology, taking measure data and harvested heads from thousands of Echinacea angustifolia plants in plots with “common garden” experimental designs. These experimental plots are located in prairie remnants, restorations, and abandoned agricultural fields that are managed as grassland habitat. Currently, the Echinacea Project has 10 established experimental plots. Some plots have multiple ongoing experiments within.
In the past few years, we have scaled back significantly on taking phenology records in the experimental plots. During 2023 and 2024, our primary goals with phenology were to a) map out the positions of flowering plants/heads within the plots, b) deploy twist ties to all flowering heads to ease measuring and harvesting, and c) to record the day of first flowering for all heads in plots to continue the long-term data collection in a more scaled-back fashion. As a result, we conducted only a few rounds of phenology per plot and did not capture the full range of flowering dates for every head. Phenology info is briefly reported on in each plot’s update along with the location of the data. This applies to the hybrid experimental plots as well.
Crew members Wyatt and Emma search can’t find positions in exPt01
Experimental plot 1 was first planted in 1996 (cleverly termed the 1996 cohort), and has been planted with nine other experiments in subsequent years, with the most recent planting being Amy Waananen’s inter-remnant crosses. It is the largest of the experimental plots, with over 10,000 planted positions; experiments in the plot include testing fitness differences between remnants (1996, 1997, 1999), quantifying effects of inbreeding (inb1, inb2), and assessing quantitative genetic variation (qGen1). It also houses a number of smaller experiments, including fitness of Hesperostipa spartea, aphid addition and exclusion, and pollen addition and exclusion (these experiments have separate posts).
In 2024, we conducted phenology in this plot between July 8th and July 18th. During measure, we visited 3123 of the 10,992 positions planted and found 2728 living plants. 83 plants were classified as “flowering” in exPt01 this year, totally 96 heads. This is a significantly fewer plants than flowered in summer 2023 (560). In summer 2024, we harvested 82 total Echinacea heads in exPt01 (including many from the ever-productive 99 south garden).
ExPt01 is also the only plot to have staples marking positions where plants that have died used to be. We added 67 staples to the experimental plot this year, but only in locations that we couldn’t find staples during measure that were already supposed to be there. We didn’t have time to get to every position with a missing staple (see where we covered here: Dropbox/CGData/125_measure/measure2024/staple2024/2024addStaplesExPt01.pdf). We did not have time to re-search locations that we called plants “can’t finds” at three years in a row in 2023 and 2024. Once these locations receive their final search, hopefully in 2025, we can put staples at them as well.
This experiment was started in 2020 by Amy Waananen to understand how the distance between plants in space and in their timing of flowering influences the fitness of their offspring. If plants that are located close together or flower at the same time are closely related, their offspring might be more closely related and inbred, and have lower fitness than plants that are far apart and/or flower more asynchronously. Plants in this experiment resulted from interremnant hand-crossings from 9 remnants: On27, SGC, GC, NGC, EELR, KJ, NNWLF, NWLF, LF. Crossing took place in 2020 and 2021, and individuals were planted in 2020 (as seed) and 2022 (as plugs). Surviving plants were assigned cgPlaIds in 2023 and incorporated into the p1 workflow. Mortality in this experiment has been high, with 80.3% of positions searched in 2024 resulting in “can’t finds.”. We did not re-search “can’t find” positions in 2024 due to time constraints (with Amy’s approval).
The inb1 experiment investigates the relationship between inbreeding level and fitness in Echinacea angustifolia. Each plant in experiment inb1 originates from one of three cross types, depending on the relatedness of the parents: between maternal half siblings; between plants from the same remnant, but not sharing a maternal or paternal parent; and between individuals from different remnants. All individuals were planted in 2001. We continued to measure fitness and flowering phenology in these plants.
The inb2 experiment investigates the relationship between inbreeding level and fitness in Echinacea angustifolia. Each plant in experiment inb2 originates from one of three cross types, depending on the relatedness of the parents: between maternal half siblings; between plants from the same remnant, but not sharing a maternal or paternal parent; and between individuals from different remnants. All individuals were planted in 2006. We continued to measure fitness and flowering phenology in these plants. In October 2024, former team member Riley Thoen recently published a paper in the Journal of Hereditary on the conservation value of small remnants using results from this experiment.
The qGen1 (quantitative genetics, or just qGen) experiment in p1 was designed to quantify the heritability of traits in Echinacea angustifolia. We are especially interested in Darwinian fitness. Could fitness be heritable? During the summer of 2002 we crossed plants from the 1996 & 1997 cohorts of exPt01. We harvested heads, dissected achenes, and germinated seeds over the winter. In the spring of 2003 we planted the resulting 4468 seedlings (this great number gave rise to this experiment’s nickname “big batch”).
data in cgData repo: ~/cgData/summer2024/exPt01Phenology
Measure data (status, size, etc.)
data in SQL database
Harvest data (IDs of harvested heads, missing achenes, etc)
detailed data in dropbox: dropbox/CGData/140_reconcile/reconcile2024/reconcileOut/2024harvestListReconciledExport.csv
data in SQL database
data in echinaceaLab package (hh.2024)
Samples collected:
82 heads harvested
At cbg for processing (counted, ready to randomize)
Products:
Publications
Thoen, R. D., A. Southgate, G. Kiefer, R.G. Shaw, S. Wagenius, The conservation value of small population remnants: Variability in inbreeding depression and heterosis of a perennial herb, the narrow-leaved purple coneflower (Echinacea angustifolia). 2024. Journal of Heredity esae055. https://doi.org/10.1093/jhered/esae055.
Page, M. L., Ison, J. L., Bewley, A. L., Holsinger, K. M., Kaul, A. D., Koch, K. E., Kolis, K. M., and Wagenius, S. 2019. Pollinator effectiveness in a composite: A specialist bee pollinates more florets but does not move pollen farther than other visitors. American Journal of Botany 106: 1487–1498. PDF
Waananen, A., G. Kiefer, J. L. Ison, and S. Wagenius. 2018. Mating opportunity increases with synchrony of flowering among years more than synchrony within years in a nonmasting perennial. The American Naturalist 192: 379-388. PDF | Appendix | online version
Muller, K. and S. Wagenius. 2016. Echinacea angustifolia and its specialist ant-tended aphid: a multi-year study of manipulated and naturally-occurring aphid infestation. Ecological Entomology 41: 51-60. PDF | online version
Shaw, R. G., S. Wagenius and C. J. Geyer. 2015. The susceptibility of Echinacea angustifolia to a specialist aphid: eco-evolutionary perspective on genotypic variation and demographic consequences. Journal of Ecology 103: 809-818. PDF
Kittelson, P., S. Wagenius, R. Nielsen, S. Qazi, M. Howe, G. Kiefer, and R. G. Shaw. 2015. Leaf functional traits, herbivory, and genetic diversity in Echinacea: Implications for fragmented populations. Ecology 96: 1877–1886. PDF
Ison, J.L., and S. Wagenius. 2014. Both flowering time and spatial isolation affect reproduction in Echinacea angustifolia. Journal of Ecology 102: 920–929. PDF | Supplemental Material | Archived Data
Ison, J.L., S. Wagenius, D. Reitz., M.V. Ashley. 2014. Mating between Echinacea angustifolia (Asteraceae) individuals increases with their flowering synchrony and spatial proximity. American Journal of Botany 101: 180-189. PDF
Ridley CE, Hangelbroek HH, Wagenius S, Stanton-Geddes J, Shaw RG, 2011. The effect of plant inbreeding and stoichiometry on interactions with herbivores in nature: Echinacea angustifolia and its specialist aphid. PLoS ONE 6(9): e24762. http://dx.plos.org/10.1371/journal.pone.0024762
Wagenius, S., H. H. Hangelbroek, C. E. Ridley, and R. G. Shaw. 2010. Biparental inbreeding and interremnant mating in a perennial prairie plant: fitness consequences for progeny in their first eight years. Evolution 64: 761-771. Abstract | PDF
Ruth G. Shaw, Charles J. Geyer, Stuart Wagenius, Helen H. Hangelbroek, and Julie R. Etterson. 2008. Unifying life-history analyses for inference of fitness and population growth. American Naturalist 172: E35 – E47. Abstract | PDF | Supplemental Material
Geyer, C.J., S. Wagenius, and R.G. Shaw. 2007. Aster models for life history analysis. Biometrika 94: 415-426. PDF | Supplemental Material
Grad student work
Drake Mullett’s PhD dissertation (2025)
Wyatt Mosiman’s MS thesis (2024)
Amy Waananen’s paper “The fitness effects of outcrossing distance depend on parental flowering phenology in fragmented populations of a tallgrass prairie forb” (with co-authors Ison, Wagenius, and Shaw) was just accepted by New Phytologist–it includes data from parents in exPt01 and progeny in exPt02.
AKA the heritability of flowering time experiment, exPt02 was designed to examine the role flowering phenology plays in the reproduction of Echinacea angustifolia. Jennifer Ison planted this plot in 2006 with 3,961 individuals selected for extreme (early or late) flowering timing, known as phenology. Using this phenological data, we explore how flowering phenology influences reproductive fitness and estimate the heritability of flowering time in E. angustifolia. In the summer of 2024, we conducted phenology between July 10th and July 19th. During measure, we visited 1,725 positions of the 3,961 positions originally planted. We measured 1,190 living plants, of which 302 were flowering with a total of 402 flowering heads (count excludes vertical developments). In the fall, we harvested 375 heads from exPt02. We observed much lower levels of seed predation by ground squirrels this year than the past few years.
data in cgData repo: ~/cgData/summer2024/exPt02Phenology
Measure data (status, size, etc.)
data in SQL database
Harvest data (IDs of harvested heads, missing achenes, etc)
detailed data in dropbox: dropbox/CGData/140_reconcile/reconcile2024/reconcileOut/2024harvestListReconciledExport.csv
data in SQL database
data in echinaceaLab package (hh.2024)
Samples collected:
375 heads harvested
at CBG for processing
Products:
Papers
Pearson, A.E., Z. Zelman, L.A. Hill, M.A. Stevens, E.X. Jackson, M.M.N. Incarnato, R.M. Johnson, S. Wagenius, and J.L. Ison. 2023. Pollinators differ in their contribution to the male fitness of a self-incompatible composite. American Journal of Botany 110(6): e16190. https://doi.org/10.1002/ajb2.16190
Reed, W. J., J. L. Ison, A. Waananen, F. H. Shaw, S. Wagenius, R. G. Shaw. 2022. Genetic variation in reproductive timing in a long-lived herbaceous perennial. American Journal of Botany 109(11) 1861–1874: https://doi.org/10.1002/ajb2.16072
Page, M. L., Ison, J. L., Bewley, A. L., Holsinger, K. M., Kaul, A. D., Koch, K. E., Kolis, K. M., and Wagenius, S. 2019. Pollinator effectiveness in a composite: A specialist bee pollinates more florets but does not move pollen farther than other visitors. American Journal of Botany 106: 1487–1498. PDF
Grad student work
Wyatt Mosiman’s MS thesis (2024)
Amy Waananen’s paper “The fitness effects of outcrossing distance depend on parental flowering phenology in fragmented populations of a tallgrass prairie forb” (with co-authors Ison, Wagenius, and Shaw) was just accepted by New Phytologist–it includes data from parents in exPt01 and progeny in exPt02.
We may have missed some other products.
exPt05:
The only experimental plot at Staffanson Prairie Preserve (SPP), exPt05 was planted to compare progeny of maternal plants from burned and unburned sections of SPP. There were originally 2800 individuals planted, but high mortality made it impractical to visit the plot row-by-row. Now, we treat the plot like demography. We use our survey-grade GPS to find plants in exPt05 that have previously flowered and add more plants to the stake file if new plants in the plot flower. In 2024, we found 17 living plants in exPt05 during flowering/total demo, 8 of which were flowering! We also observed (but did not take data on) additional basal plants within the plot boundaries that appeared to be growing on a 1×1 meter grid. ExPt05 persists!
Team Echinacea established quantitative genetics experiments to quantify additive genetic variance of fitness in Echinacea, with the idea that we can estimate evolutionary potential of study populations. The plants in qGen2 and qGen3 are plants in the 1996, 1997, and 1999 cohorts. These plants were crossed with pollen from plants in remnants to produce seed for qGen2 and qGen3, which now inhabit exPt08. Originally, 12,813 seeds were sown in the common garden. Seeds from the same cross (shared maternal and paternal plants) were sown in meter-long segments between nails. In the summer of 2024, we conducted phenology between July 10th and July 19th. A total of 3,253 seedlings were originally found, but due to gradual mortality we only searched 448 positions in 2024 for plants, and we found evidence of 314 living individuals. We identified 24 flowering plants in with a total of 29 heads, of which we harvested 23.
data in cgData repo: ~/cgData/summer2024/exPt08Phenology
Measure data (status, size, etc.)
data in SQL database
Harvest data (IDs of harvested heads, missing achenes, etc)
detailed data in dropbox: dropbox/CGData/140_reconcile/reconcile2024/reconcileOut/2024harvestListReconciledExport.csv
data in SQL database
data in echinaceaLab package (hh.2024)
Samples collected:
23 heads harvested
at CBG for processing
Products:
None… yet!
tplot
tplot is located within the bounds of exPt08. Plants of many species here were rescued from the landfill site. They we transferred as chunks of prairie sod, and individual transplants. This year during flowering and total demo, we encountered 14 living Echinacea plants from which we harvest 12 heads.
Demography data: head counts, rosette counts, etc.
demap input files have been updated with 2024
Spatial location for all flowering and some basal (total demo) plants
demap input files have been updated with 2024
Harvest data (IDs of harvested heads, missing achenes, etc)
detailed data in dropbox: dropbox/CGData/140_reconcile/reconcile2024/reconcileOut/2024harvestListReconciledExport.csv
data in echinaceaLab package (hh.2024)
Samples collected:
12 heads harvested
at CBG for processing
Products:
None… yet!
Experimental plot management:
Just like other areas of the prairie, our experimental plots need management! Here’s a list of the stewardship activities that we conducted in or for our plots during 2024:
NU MS student Maddie Sadler shows off our sweet clover haul
None of our experimental plots burned in the fall or spring prior to the 2024 growing season
Collected seed to plant in p1, p2, & p8, including:
1) Elise collected Carex brevior, Carex bicknellii, and Carex gravida from several sites and from plants that we established a few years ago near exPt01
2) Liam established Viola pedatifida production tub,
3) We collected a few additional species (including Galium boreale, Solidago missouriensis, Astragalus adsurgens, Dichanthelium leibergii, and Bromus kalmii) we plan to establish via plug in spring 2025
Psst – next year person writing this report; I ran my numbers using the script Dropbox/echProjAdmin/projectStatusReports/psr2024/wmGatherDataForReports2024.R. Maybe this can help you out.
We’re interested in investigating what resources are available to Echinacea visitors and learning more about the pollen and nectar Echinacea produces. We hope to learn if the nutritional resources available differ before and after burns. In 2022, Britney House developed methods for collecting nectar from Echinacea using microcapillary tubes. Read more about her methods here.
During the summer of 2024, the team collected pollen and nectar samples from Echinacea angustifolia at 12 sites in and around Solem Township, MN. We searched for and shot the ~10 plants (or, if few were available, as many as we could find) at each site that were closest to a random point. We then bagged up to five of the heads with pollinator exclusion bags for those 10 plants. Throughout the duration of their flowering, we collected pollen from all bagged plants and nectar from five of them per site.
We removed bags from pollen/nectar plants and backup plants when they were done flowering, and we collected until a a limit was set of a cumulative 10mm of nectar from each plant. This year we also collected immature florets from each bagged head at the start of sampling.
Following some experimentation, we conducted nectar collection only in the afternoons, while pollen collection could be done any time of day. In total, we collected vials of pollen, nectar, and vials containing immature florets from 60 plants. Pollen and nectar tubes were given to Rahul Roy at St. Catherine University in St. Paul, who will be doing pollen and nectar analysis. Tubes containing immature florets were sent with Grace Hirzel at North Dakota State University in Fargo, ND for pollen grain count and size analysis.
Data entry for collection datasheets is ongoing. Pollen data entry is started and nectar data is a little over half done with the help of Emma Reineke. Emma will also be using part of this dataset for her senior thesis project at the University of Minnesota.
Scans can be found at: Dropbox/teamEchinacea2024/z.pollenNectarDataEntry/scans.
Start year: 2024
Location: Various prairie remnants around Solem Township, MN
Team members involved with this project: Summer team 2024, Rahul Roy (St. Kate’s), Emma Reineke (University of Minnesota), Jarrad Pasifrika and Grace Hirzel (North Dakota State University)
Products: pending
Funding: ENRTF
Grace Hirzel taking off a pollen excluder bag on an Echinacea angustifolia plant.
Echinacea pallida is a species of Echinacea that is not native to Minnesota. It was mistakenly introduced to our study area during a restoration of Hegg Lake WMA around 2006 (Stuart’s recollection). This is concerning, because we don’t know how a similar species may impact or local Echinacea angustifolia! Will they hybridize? Could pallida outcompete angustifolia? Ever since pallida have started springing up, Team Echinacea has visited the pallida restoration, taken flowering phenology, and collected demography on the non-native plant. We have decapitated all flowering E. pallida each year to avoid cross-pollination with the local Echinacea angustifolia. Each year, we record the number of heads on each plant and the number of rosettes, collect precise GPS points for each individual, and cut off all the heads before they produce fruits.
This year, we cut E. pallida heads on June 26th. Overall, we found and shot 172 flowering E. pallida plants with 512 normal heads in total, averaging 2.98 heads per plant, though the max was 20 on a single plant! These non-native plants were hearty with an average rosette count of 7.20 rosettes and an astounding individual with a maximum of 88 rosettes. We did not take phenology data on E. pallida this year.
Team 2024 returns to their vehicles after flagging, taking demography data on, and decapitating Echinacea pallida at Hegg Lake WMA
Demography data: head counts, rosette counts, etc.
data in aiisummer2024 repo: ~/aiisummer2024/demo/demoGood2024.txt
Spatial location for every flowering E. pallida
data in aiisummer2024 repo: ~/aiisummer2024/surv/survGood2024.txt
Samples collected:
Echinaceapallida were not collected: decapitated heads were left on the ground next to the plants
A flowering echinacea at the aptly named nearby remnant “near pal” looked suspiciously like a hybrid (more robust than an angustifolia). We put a pollinator exclusion bag on the single head to prevent pollen spread, and later harvested the head and brought it back to the lab, where it is currently in the seed dryer. Keep an eye on tag 29239 in the future
Products:
None… yet! Besides a prairie with significantly less E. pallida reproduction
You can find more information about E. pallida flowering phenology and previous flog posts on the background page for the experiment.
Experimental plot 6 was the first E. angustifolia x E. pallida hybrid plot planted by Team Echinacea. A total of 66 Echinacea hybrids were originally planted. All individuals have E. angustifolia dams and E. pallida sires. In 2024, we visited 23 positions, 4 of which were can’t find year 3 in 2023 and didn’t get their final double check. We found living plants at all positions but those four (so, 19)! Last year, for the first time, 3 plants flowered in this plot. This year, no plants flowered.
Start year: 2011 (crossing) and 2012 (planting)
Location: Wagenius property
Overlaps with:
Common garden experiment
Data collected:
Measure data (status, size, etc.)
data in SQL database
Samples collected:
None (no flowering plants)
Products:
None… yet!
You can find more information about experimental plot 6 and previous flog posts about it on the background page for the experiment.
Experimental plot 7 is the second E. pallida x E. angustifolia plot. It contains conspecific crosses of each species as well as reciprocal hybrids, totaling 294 pdeigreed individuals. We took phenology records between July 10th and July 18th. There were 42 flowering plants this year; from these we harvested 87 heads. Heads in this plot were covered by pollinator exclusion bags during the growing season to prevent cross-pollination with nearby Echinacea populations.
Stuart demonstrates proper measuring technique in exPt07
Start year: 2012 (crossing) and 2013 (planting)
Location: Hegg Lake WMA (MN DNR)
Overlaps with:
Common garden experiment
Data collected:
Phenology data (dates of flowering stages)
data in cgData repo: ~/cgData/summer2024/exPt79Phenology
Measure data (status, size, etc.)
data in SQL database
Harvest data (IDs of harvested heads, missing achenes, etc)
detailed data in dropbox: dropbox/CGData/140_reconcile/reconcile2024/reconcileOut/2024harvestListReconciledExport.csv
data in echinaceaLab package (hh.2024)
Samples collected:
87 heads harvested
at CBG for processing
Products:
None… yet!
You can find more information about experimental plot 7 and previous flog posts about it on the background page for the experiment.
There were originally 745 seedlings planted in exPt09. Experimental plot 9 is a hybrid plot, but, unlike the other two hybrid plots, we do not have a perfect pedigree of the plants. That is because the E. angustifolia and E. pallida maternal plants used to generate seedlings for exPt09 were open-pollinated. At this point, some but not all plants in this plot were tested for paternity, revealing that there are some hybrids. This year, we took phenology records between July 9th and July 18th. During measuring, we searched at 292 positions and found evidence of 234 living plants in 2024. Of these individuals, 70 were flowering. We harvested 110 heads from this plot! Heads in this plot were covered by pollinator exclusion bags during the growing season to prevent cross-pollination with nearby Echinacea populations.
The team runs out reel tapes to aid in measuring exPt09
Start year: 2014
Location: Hegg Lake WMA (MN DNR)
Overlaps with:
Common garden experiment
Data collected:
Phenology data (dates of flowering stages)
data in cgData repo: ~/cgData/summer2024/exPt79Phenology
Measure data (status, size, etc.)
data in SQL database
Harvest data (IDs of harvested heads, missing achenes, etc)
detailed data in dropbox: dropbox/CGData/140_reconcile/reconcile2024/reconcileOut/2024harvestListReconciledExport.csv
data in echinaceaLab package (hh.2024)
Samples collected:
110 heads harvested
at CBG for processing
Products:
None… yet!
You can find out more information about experimental plot 9 and flog posts mentioning the experiment on the background page for the experiment.
During the summer of 2024, Team Echinacea completed the second year of its ENRTF funded project to better understand how prescribed fire influences ground nesting bee habitat, food resources, and diversity. Understanding the associations between land management methods and ground nesting bees is essential for providing reccomendations to policymakers and practitioners interested in native bee conservation.
We surveyed solitary bee diversity and nesting habitat before and after prescribed fires in a subset of 30 prairie remnants and 15 prairie restorations to determine how prescribed fire affects solitary bee nesting habitat and abundance. We used emergence traps to sample the community of solitary ground nesting bees. This was complemented by detailed measures of soil and litter to characterize how prescribed burning influences the nesting habitat (read more here).
2024 REU student Zach Zarling deploys an emergence trap at a site near Hoffman, Minnesota
We deployed emergence traps at our random “burn and bee points”(BBPTs) in prairie remnants and restorations from early June to mid September. Our deployments spanned three rotations (4-6) of BBPTs and we put out a total of ~1,159 emergence traps. On reccomendation from Dr. Alex Harmon-Threatt, we also performed 10 minute “pollard walks” on deployment to estimate the number of foraging bees at each site. These foraging numbers will be compared to nesting incidence as part of Ian Roberts’ thesis project.
As of December 21st, specimens caught in this year’s deployments have been pinned, labeled, and transported from Chicago Botanic Garden to the University of Minnesota, where Zach Portman, a bee taxonomist, will identify them. Team Echinacea also collected lots of non-bee bycatch while processing specimens collected in the traps: including millipedes, flies, and even a prairie skink! To avoid wasting these specimens, we plan to categorize this bycatch into broad taxonomic groups (like Dipterans, Orthopterans, etc) and examine potential associations between our experimental treatments and general arthropod diversity across our study sites.
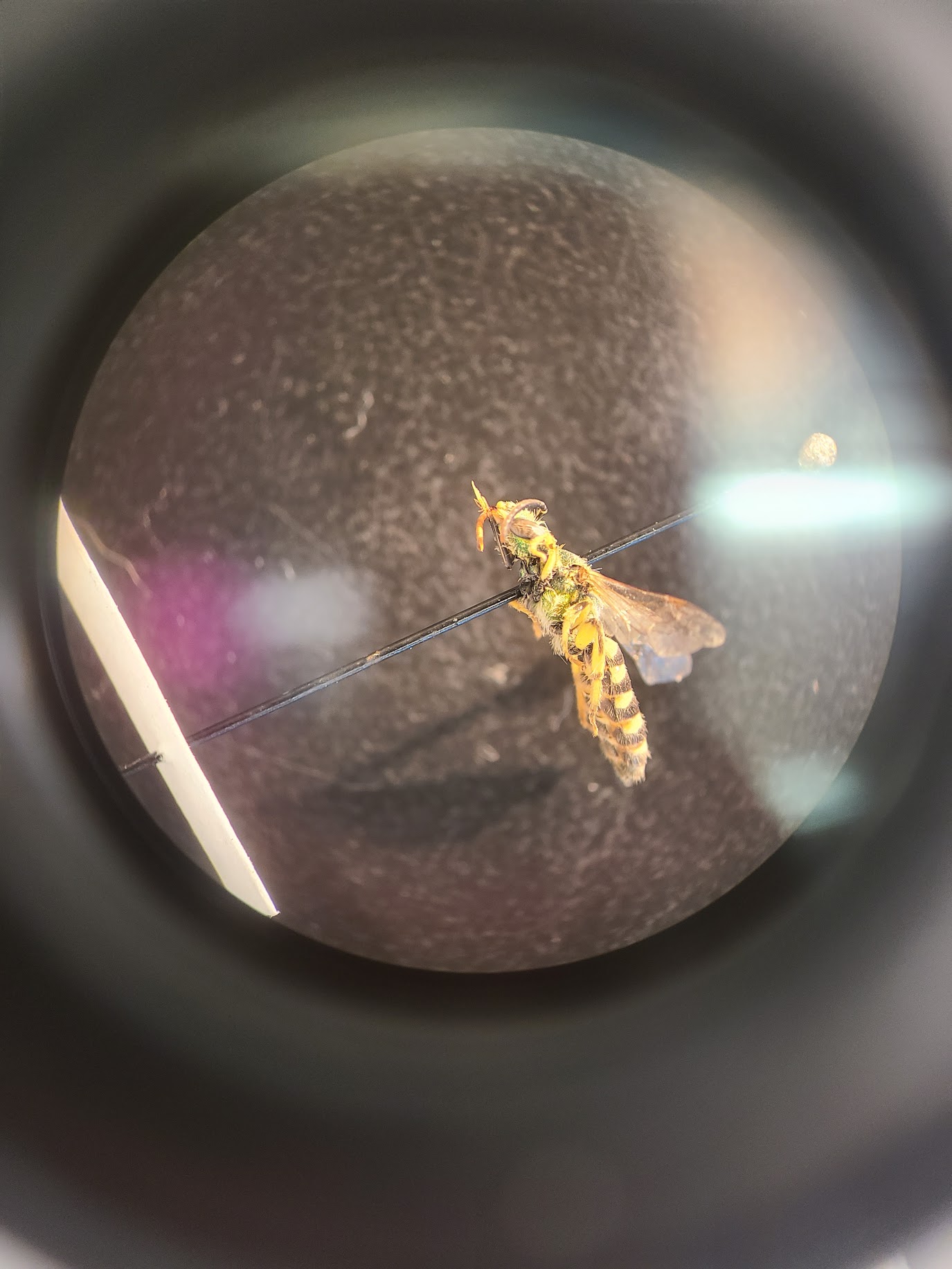
Pinned specimen from 2024’s emergence trapping, likely an Agapostemon virescens.
While working on pinning and processing specimens, Ian Roberts produced a poster containing analyses from the 2023 emergence trapping data to present at Entomology 2024. The poster can be viewed here. Future data analyses will feature data from both sampling years, as well as microhabitat measurments and and diversity indices.
Start year: 2023
Location: prairie remnants and restorations in Solem Township, MN.
Data collected: insect samples, counts of foraging bees
Samples or specimens collected: Pinned bees are currently being identified at University of Minnesota. Bycatch is in the freezer at Chicago Botanic Garden.
Products: poster presented at Entomology 2024 (see above for link)
During summer 2024, Team Echinacea continued to collect data on local environmental conditions in order to understand which environmental factors are associated with good habitat for ground-nesting bees. These data complement emergence trapping for our ENRTF funded research on fire’s influence on ground nesting bees habitats. We sampled local environmental conditions near randomly placed “burn and bee points” (BBPTs) in prairie remnants and restorations.
A deployed emergence trap next to a set of marking flags. Microhabitat data was sampled within a meter of these marking flags.
Unlike the 2023 season, we did not collect data on light levels at BBPTs this year. Instead, we measured soil temperature just under the surface using a digital thermometer, along with soil compaction via a penetrometer and litter depth via a meter stick.
Team Echinacea conducted microhabitat assessments for remnant prairies at rotation 4 BBPTs. Over the summer, we took microhabitat assessment measurements at a total of 241 measurments.
Start year: 2024
Location: prairie remnants and restorations in Solem Township, MN
The Viola propagation tub Team Echinacea installed this past summer was designed with a few goals in mind. First of all, we wanted the tub to produce Viola pedatifida seed that could be used to help seed this important spring native throughout our remnants and experimental plots. We have already seen a pretty good return on our investment from the seed production end, just this fall 10 ripe pods have been harvested each containing between 20 and 30 seeds so this season around 250 seeds have been collected. Additionally, 9 Immature pods were collected before winterizing the plot, so hopefully, some good seeds will come from those as well. The second big goal we wanted to explore with this setup is learning more about small-scale production plots. Several other species have been known to grow well in beds like violets and hopefully, through managing this violet plot, we can learn more about what to expect. Other notable species that may go into future production plots are Ranunculus rhomboideus, Fragaria virginiana, Geum triflorum, and many others. Other good data to collect may come from the survivability of plants directly transferred from remnants and data on germination and survivability of seedlings.
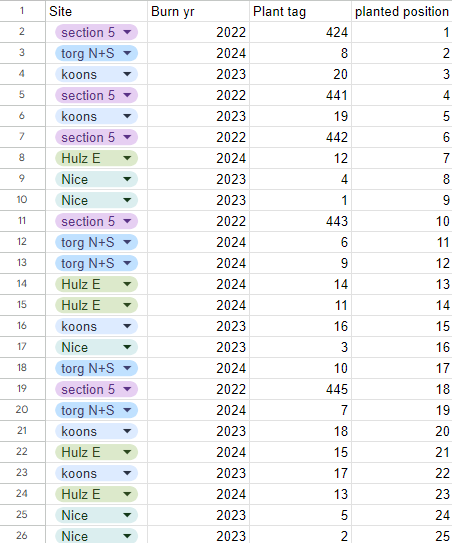
Plot Layout and Plant Data
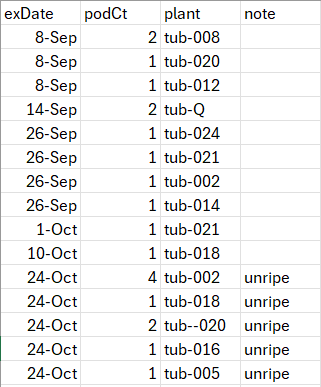
The plot is laid out below with the corresponding position numbers. The left spreadsheet lays out the ID and source for each plant, and the right contains data on pods collected so far.
Collecting Pods
Throughout the fall, 2 pods were followed from flowering to splitting of the pod. Pictures were taken each day along the entire course each morning before lunch. each pod took 17 days from flower to splitting open. Photos here will be arranged in inverse chronological order starting with pod splitting and then counting backwards. Moving forward we would like to learn what the exact features are of a pod that is ready to harvest. So far these photos have shown 2 likely indicators. 1) the pod starts very green and small and slowly turns a pale cream color and fills out as it matures. 2) The pod stem transitions from a bent gooseneck shape into a more upright position while lightening in color. When a pod is ready the stem tends to be fairly straight or has a soft curvature.
Violet 8
Violet 11
Tucking in for Winter
Oct 24 2024 the violets seed production was slowing down and with winter weather on its way it was decided that we should tuck the violets in and cover them. Firstly litter and any seeds visible on the surface of the tub. Next, a 4-inch mulch of Straw cut from P1 was applied across the entire surface of the tub, being sure to cover all green stems from the violas. Lastly, a Sheet of hardware cloth was cut to the length of the tub to prevent the intrusion of digging animals. This sheet was bent around the edges of the tub and then weighed down by several stones. Afterwards, the violas should be safe until next summer.
Spring 2025
A few things will definitely be on the docket for next spring. First off after the snow recedes the wire and mulch should be removed as soon as possible to ensure the soil warms and the volets get enough sunlight. As soon as the soil is removed an inch or so of compost should be applied across the entire bed. Once the violets emerge then the mulch can be reapplied to prevent weed development. Flowers should develop quickly and the job of picking ripe pods will almost certainly last all summer.
The aphid addition and exclusion experiment was started in 2011 by Katherine Muller. The original experiment included 100 plants selected from exPt01 that were each assigned to have aphids either added or excluded across multiple years. The intention is to assess the impact of the specialist herbivore Aphis echinaceae on Echinacea fitness.
In 2024, 41 of the original 100 plants were alive, two of which flowered. However, we did not see any aphids anywhere while measuring exPt01. We have not conducted fieldwork for this experiment since 2022, when team members Emma Reineke and Kennedy Porter were in charge of the experiment and did not find any aphids in exPt01, so they introduced a new population of Aphid echinaeceae into the plot. Learn more in the 2022 summer aphid update.
Aphids Wyatt Mosiman saw in exPt02 this summer. Wrong plot, aphids!
Plant survival and measurements were recorded as part of our annual surveys in P1 and eventually will be found in our SQL database.
Samples collected:
2 heads from plants included in this experiment are at the Chicago Botanic Garden awaiting processing: AD-1728 and AZ-1744 in the exPt01 2024 batch
Products:
Andy Hoyt’s poster presented at the Fall 2018 Research Symposium at Carleton College
2016 paper by Katherine Muller and Stuart Wagenius on aphids and foliar herbivory damage on Echinacea
2015 paper by Ruth Shaw and Stuart Wagenius on fitness and demographic consequences of aphid loads
You can read more about the aphid addition and exclusion experiment, as well as links to prior flog entries mentioning the experiment, on the background page for this experiment.
Reproduction in plants can be limited by access to pollen and resources. We previously found that Echinacea plants in the remnants are pollen limited, meaning that if they had access to more pollen, they would produce more seeds. However, the long-term effects of pollen limitation are unknown. Do plants that are super pollen saturated and have high amounts of pollen have a higher lifetime fitness than plants that are pollen limited? Also, we know that the plants in the remnants are pollen limited, but are the plants in the common garden environment also pollen limited? To answer these questions and more, 13 years ago Gretel randomly selected 39 plants from p1; half of these plants were randomly assigned to the pollen addition group, and the others were assigned to pollen exclusion. Every year, plants in the pollen exclusion have their heads bagged and they are not pollinated, while we hand cross every style in the pollen addition group. An additional 53 plants in p1 were selected to be part of a control group where pollen was neither added nor excluded.
In the summer of 2024, NONE of the original 39 addition/exclusion plants were flowering. If any had been flowering, the exclusion treatment plants would be covered with exclusion bags to prevent pollination, and the addition plants would be hand-pollinated multiple times throughout the summer. One plant in the control group produced a single flowering head.
Collaborator and NDSU postdoc Grace Hirzel places pollinator exclusions bags on Echinacea heads in the remnants. If any of the plants in the polLim experiment from the non-control group had flowered this year, it would’ve looked like this!
You can find more information about the pollen addition and exclusion experiment and links to previous flog posts regarding this experiment at the background page for the experiment.
It has been a packed few days here in Phoenix, and I’d like to describe some of the presentations I saw at yesterday’s ground nesting bee symposium. There was a broad range of topics, including the microbe communities found on pollen balls, brood parasite and host interactions, rare desert bees, and more! Nick Dorian had great results to share from his mark-recapture work in New Jersey, which was great to see after he demonstrated that method to Team Echinacea over the summer. Lily Fulton’s presentation really stood out to me: She is also studying prescribed fire, at a long term experimental fire site in Florida longleaf pine flatwoods. they found higher bee and wasp abundances during burn years, as well as higher proportions of bare ground. This is strikingly similar to our research findings! It was great to chat with her about it over my own poster.
Title slide of Lily Fulton’s presentation. My poster! It is showing the first year of data from the ENRTF project, since we haven’t sent our 2024 bees to Zach yet.
It’s been incredible getting to meet so many researchers who care about native bees. Many stopped by to have a look at the poster! I’ve cited many of them in my previous work, so running into them in person is almost like meeting a celebrity. One consistent interest I’ve gotten from attendees is to look a the community composition of bees in the different treatment groups. Perhaps the increased abundance in the year of a burn we’re seeing is because of a cohort of burn-loving ground nesters, who don’t stick around in following years. It’ll be exciting to look into!
Shot of the poster sessions. There were a ton of applications, so space was pretty limited.
Overall, this has been a fantastic experience and opportunity to expand my network. I got to reconnect with some old friends, and make plenty of new ones. I can’t wait to get back to work on my thesis, now that I know who might be viewing it once it’s published! Expect more on that in the next few months.
As I’m preparing to write updates on experiments for 2024, it was brought to my attention that we don’t have one post summarizing all our updates from summer 2023! So, while you eagerly await this year’s news, enjoy a refreshing blast from the past. Here’s what we did last summer!
Members of Team Echinacea 2023 measure plants in our hybrid experimental plots at Hegg Lake WMA























































{kind=link}