During the summer of 2020, I designed an observational study to assess the extent to which reproduction in self-incompatible species is limited by spatial proximity to mates. While many researchers assume mate-limited reproduction is common in prairie species, few studies demonstrate this relationship in species other than Echinacea. I hypothesize that many self-incompatible prairie species produce few viable seeds because of small population size and spatial isolation from mates. To test this hypothesis, I selected 8 common self-incompatible native prairie species and 25 remnant sites within which to work. I quantified isolation in meters from the two nearest flowering conspecifics and collected seeds for more than 1,100 individual plants at 25 prairie remnant sites. I hope to complete lab work over the next 6 months where I will be cleaning and counting viable seeds for each individual plant. Results from this study will provide evidence about how widespread mate-limited reproduction is in self-incompatible prairie species.
Start year: 2020
Location: Douglas County, Minnesota; 25 roadside remnant sites Information about how many species of each species were sampled from each site, check out this file ~Dropbox/teamEchinacea2020/leaRichardson/siteCheckList8.29.20.xlsx
Materials collected: Seed heads collected from over 1,100 plants will be stored at the Chicago Botanic Garden but are currently in my Evanston apartment.
Data collected: Find data related to this project in the aiisummer2020 repository in ~aiisummer2020/plasInRems/leaStuff/
Throughout the summer, I designed and collected materials to establish an experiment in experimental plot 1 to study parasites and their impact on the community of host plants they live in. Parasitic plants are plants which absorb nutrients from neighboring plants. Parasitism is an important part of nutrient cycling in many ecosystems and parasite scientists hypothesize it to be an important part of prairie ecosystem maintenance.
This experiment has three factors, each with two levels (presence or absence), but three factor-level combinations are impossible, because the presence of parasites is confounded with presence of soil. Which translates to me having 216 row x position combinations in which I randomly assigned Comandra umbellata, Pedicularis canadensis, and soil plugs. However, because roots trap soil and therefore soil is always carried in with parasites, the two are confounded and so we used soil transplants to account for this.
I developed this experiment to address questions about the impact native parasitic plants have on plant community members. In late October I harvested biomass from my pipp places to understand how species diversity and abundance change after planting parasites.
Start year: 2019
Location: Douglas County, Minnesota; exPt 1
Overlaps with: Experimental plot management, Hesperostipa common garden experiment
Materials collected: 216 .1 x 1m strips of dried biomass are stored at the Chicago Botanic Garden.
Data collected: Find data related to this project including the planting scheme in the cgdata repository in ~cgdata\summer2019\Hemiparasites (note/the key for HemiparaMap: C. umbellata = Blue, P. canadensis = Red, C + P = Purple, Soil plugs = Brown, Just seeds = Green).
Team Echinacea continued the aphid addition and exclusion experiment started in 2011 by Katherine Muller. The original experiment included 100 plants selected from exPt01 which were each assigned to have aphids either added or excluded through multiple years. The intention is to assess the impact of the specialist herbivore Aphis echinaceae on Echinacea fitness.
The 2020 aphid team was Anna Allen and Allie Radin. They located 25 living exclusion plants and 16 living addition plants. The experiment was conducted from July 6th to August 19th, with the final visit consisting only of observation. Aphids were moved only during four visits from late July to mid-August due to late arrival and low numbers of aphids. Only one or two aphids were applied to each plant during each visit. They recorded the number of aphids present in classes of 0, 1, 2-10, 11-80, and >80. They also recorded the number of aphids added.
Aphids on an Echinacea leaf
Start year: 2011 Location: Experimental Plot 1 Overlaps with: Phenology and fitness in P1 Data collected: Scanned datasheets are located at ~Dropbox\teamEchinacea2020\allisonRadin\aphidAddEx2020.
Products:
Andy Hoyt’s poster presented at the Fall 2018 Research Symposium at Carleton College
2016 paper by Katherine Muller and Stuart on aphids and foliar herbivory damage on Echinacea
2015 paper by Ruth Shaw and Stuart on fitness and demographic consequences of aphid loads
You can read more about the aphid addition and exclusion experiment, as well as links to prior flog entries mentioning the experiment, on the background page for this experiment.
Supplemental pollen — pollen that an Echinacea head might not otherwise receive—could increase a plant’s fitness. But does this extra pollination lead to a tradeoff in survival or flowering consistency? Since 2012, we have been manipulating the amount of pollen Echinacea plants receive – either no pollen, or lots of pollen – and recording how this affects their fitness and survival. In 2012 and 2013 we identified flowering E. angustifolia plants in experimental plot 1 and randomly assigned one of two treatments to each: pollen addition or pollen exclusion. The team bagged the heads of all plants and hand-pollinated the addition treatment, and did not manipulate the exclusion plants further. Plants receive the same treatment across years.
In summer 2018, 14 of the 26 plants alive in the pollen addition and exclusion experiment flowered, producing a total of 25 heads. This year none of those plants flowered. Of the original 38 plants in this experiment, 12 of the exclusion plants and 14 of the pollen addition plants are still alive. No plants died between 2018 and 2019. This year’s data were unique among the eight years of data collected, because not a single plant in the experiment produced even a single head. The dramatic decrease in flowering rates this year may help or hinder us in analyzing this data set and providing answers to this eight-year question.
Tris did not find significant demographic differences between plants which received pollen exclusion, addition or open pollination treatments.
Start year: 2012
Location: exPt1
Physical specimens: We harvested no specimens this year
Data collected: Plants survival and measurements were recorded as part of our annual surveys in P1 and can be found with the rest of the P1 data in the R package EchinaceaLab.
Michael presented a poster on the polLim experiment at MEEC
2019, which you can find here
Tris also presented a poster on polLim at MEEC 2019, which you can find here
You can find more information about the pollen addition and
exclusion experiment and links to previous flog posts regarding this
experiment at the background page for the experiment.
Team Echinacea established quantitative genetics experiments to determine the additive genetic variance of fitness in Echinacea, with the idea that we can estimate evolutionary potential of study populations. Quantitative genetics experiments 2 and 3 (qGen2 and qGen3) represent the third generation of Echinacea in our common garden experiments. The grandparents of qGen2 and qGen3 are the 1996 and 1997 gardens. Plants from these experiments were crossed to generate qGen1 (a.k.a. Big Batch), and plants in qGen1 were crossed to produce seed for qGen2 and qGen3, which now inhabit exPt8.
We visit exPt8 every year to assess fitness of Echinacea in the plot. Originally, 12,813 seeds were sown in the common garden. Seeds from the same maternal and paternal plant were sown in meter-long segments between nails. A total of 3253 seedlings were originally found, but only 669 plants were found alive in 2019.
Jay, John, and Avery assess fitness of young Echinacea in exPt8. They’re so tiny (the Echinacea, that is… Jay, John, and Avery are regular sized).
In an exciting turn of events, we found a flowering plant in qGen2 this year! This was the first flowering plant found in exPt8. Fortunately for our one flowering plant, it had four flowering friends to cross with from the Transplant Plot. We took phenology data on the qGen2 head, measured it, and harvested it.
The presence of a flowering plant influenced Riley Thoen to make a new measuring form for exPt8 in 2020. In the past, the exPt8 measuring form was very different from other measuring forms. Through 2019, we measured all leaves of basal plants in exPt8; we only measure the longest basal leaf in other plots. Riley designed the 2020 exPt8 measuring form to mirror the measuring forms from other common gardens. In the future, the exPt8 measure form will have a head subform and team members will only have to measure the longest basal leaf of each plant found.
Start Year: 1996 and 1997 (Grand-dams), 2003 (qGen1 – dams), 2013 and 2015 (qGen2 and qGen3, respectively)
Data/material collected: phenology data on the flowering plant and transplant plot plants (available in the exPt1 phenology data frames in the cgData repo), measure data (cgData repo), and harvested heads (data available in hh.2019 in the echinaceaLab package; heads in ACE protocol at CBG).
The inbreeding 2 experiment was planted in exPt1 in 2006 to determine how genetic drift is differentially affecting average fitness of remnant populations. In 2005, team members crossed common garden plants from seven remnant populations. There are three cross types: inbred (crossed to a half-sib; I), within population (randomly chosen; W), and between population (B). Each year, team members assess flowering phenology and fitness of Echinacea in the inb2 common garden.
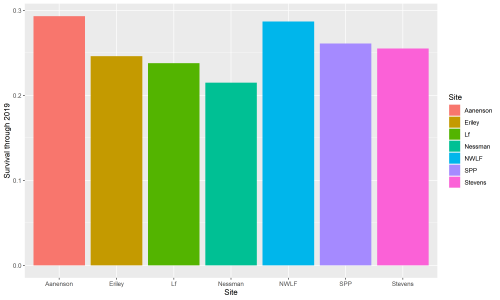
In 2019, the team searched for Echinacea at 508 positions of the original 1443 positions planted in inb2. In total, we found 351 living plants. Four plants flowered in 2019 but only three produced achenes. Since 2006, 163 Echinacea in inb2 have flowered; they have produced a total of 336 flowering heads.
This winter, Riley Thoen is working on analyzing data and drafting a manuscript for inb2. In these endeavors, he found a small discrepancy in inb2 data: not all plants that were planted in the inb2 plot have a complete pedigree. Therefore, only a subset of the total can be used for analysis. A total of 1136 plants with a complete pedigree were planted in inb2, and of those, 277 were found alive in 2019. All four plants that flowered in 2019 have known pedigrees. A total of 138 plants of known pedigree have flowered and they have produced 284 total heads since the plot was planted in 2006. Surprisingly, within-remnant crosses have the lowest survival of all cross types, at 20%. Inbred crosses have 24% survival and between-remnant crosses have 30% survival. Riley is starting to push data analysis forwards and will certainly post updates on the flog when more discoveries are made!
Data/material collected: flowering phenology on the flowering plants (available in the exPt1 phenology data frames in the cgData repo), measure data (cgData repo), and harvested heads (data available in hh.2019 in the echinaceaLab package; heads in ACE protocol at CBG).
Products:
Shaw, R. G., S. Wagenius and C. J. Geyer. 2015. The susceptibility of Echinacea angustifolia to a specialist aphid: eco-evolutionary perspective on genotypic variation and demographic consequences. Journal of Ecology 103: 809-818. PDF
Kittelson, P., S. Wagenius, R. Nielsen, S. Qazi, M. Howe, G. Kiefer, and R. G. Shaw. 2015. Leaf functional traits, herbivory, and genetic diversity in Echinacea: Implications for fragmented populations. Ecology 96: 1877–1886. PDF
Monitoring
reproductive fitness in the remnant populations is a staple of Team Echinacea’s
summer activities. Understanding the reproductive success of plants in remnant
populations provides insight to a vital demographic rate contributing to the
persistence (or decline) of remnant populations in fragmented environments.
In summer
2019, we harvested 40 seedheads to study patterns of reproductive fitness in 8
remnant Echinacea populations (ALF, EELR, KJ, NWLF, GC, NGC, SGC, NNWLF) (the
same populations used where I studied phenology and gene flow). I randomly selected 1/3 of
flowering heads at each remnant to harvest. In addition, I collected all seedheads
from especially small or isolated remnants (specifically, GC, KJ, and the
cluster of plants just north of EELR).
In early
January, I dissected the seedheads. I extracted the achenes by row so that I
will be able to observe temporal variation in seed set within heads. Ideally,
next I will x-ray the achenes and assess seed set by observing the proportion
of achenes that contain embryos. However, the x-ray machine at the Chicago
Botanic Garden is currently out of service, so instead I may need to weigh or
germinate the achenes to see if viable embryos are inside.
Extracting achenes by row, so that I know which achenes resulted from florets that flowered early (i.e., at the bottom of the seedhead) or late (i.e., at the top of the seedhead). Tedious but possible!
Start
year: 1996
Location: Roadsides,
railroad rights of way, and nature preserves in and around Solem Township, MN
You can
read more about reproductive fitness in remnants, as well as links to prior
flog entries mentioning the experiment, on the background page for this experiment.
In summer
2019, I completed a second season of field work for a study monitoring pollen
movement between remnant populations. In summer 2018, I chose two focal areas,
the NW sites in the study area (populations: ALF, EELR, KJ, NWLF, GC, SGC, NGC,
KJ, NNWLF) and SW sites (populations: LC, NRRX, RRX, YOH, and two large populations
in between these sites). This summer, I limited the study to the NW sites. As
in 2018, I mapped and collected leaf tissue from all individuals in the study
areas and harvested seedheads from a subset of these individuals (see Reproductive Fitness in Remnants). In addition, I monitored the
flowering phenology of all of the flowering plants in these populations (see
Phenology in the Remnants).
Now, I am
working on extracting and genotyping the DNA from the leaf tissue samples and a
subset of the seeds I collected. This takes a long time! I will use the
microsatellite markers that Jennifer Ison developed in her dissertation to
match up the genotypes of the offspring (i.e., the seeds) with their most
likely father (i.e., the pollen source). To analyze patterns of gene flow, I
will assess how individuals’ location and timing of flowering influence their
reproductive success and distance of pollen movement.
In
addition, last summer we planted all of the seedlings from 2018 in the
experimental plot that John Van Kempen set up at West Central Area High School.
We will continue to monitor these seedlings to understand how pollen movement
distance (or the distance between parents) influences offspring fitness.
Here is the team after we planted nearly 298 seedlings in the experimental plot at WCA!
Start
year: 2018
Location: Roadsides, railroad rights of way, and nature preserves in and around
Solem Township, MN
Products: I presented a poster based on
the locations and flowering phenology of individuals from summer 2018 at the
International Pollinator Conference in Davis, CA this summer. The poster is
linked here: https://echinaceaproject.org/international-pollinator-conference/.
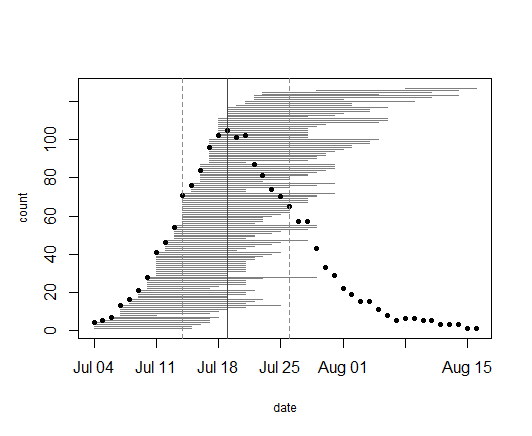
In 2019,
we collected data on the timing of flowering for 95 flowering plants (127
flowering heads) in 8 remnant populations, which ranged from 1 to 29 flowering heads.
The earliest bloomers (four plants at four different remnants) initiated
flowering on July 4. Plant 24050 in the aptly named remnant population North of
Northwest of Landfill was the latest bloomer, shedding its last anthers of
pollen on August 16. Township mowers had mowed over this plant earlier in the
season, which is perhaps why it took longer for it to sprout a new flowering
stem. Altogether, the flowering season was 43 days long. Peak flowering was on
July 19, when 105 heads were flowering.
This season
marked the 19th season of collecting phenology records in remnant populations! Though
we do not have data for all populations every year, Stuart monitored phenology
in all of our remnant populations in 1996 and in following years (2007, 2009,
2011-2019) students and interns studied phenology in particular populations. From
2014-2016, determining flowering phenology was a major focus of the summer
fieldwork, with Team Echinacea tracking phenology in all plants in all of our
remnant populations. The motivation behind this study is to understand how
timing of flowering affects the mating patterns and fitness of individuals in
natural populations.
Flowering
occurred much later this season than previous years, with peak flowering
falling a full 14 days later in the year than 2018, when flowering started on
June 20, and 10 days later than 2017. Of all the years that we data for flowering
phenology in the remnant populations in and around Solem Township, this season was
the second-latest, with only the 2013 season beginning later, on July 6.
However, this observation comes with the caveat that sampling effort varied
between years and some years focused on particular contexts, such as population
where a portion had experienced a spring burn (see Fire and Flowering at SPP). Many other plants and animals in
Minnesota seemed to have delayed phenology this spring and summer, perhaps a
result of an unusually wet and snowy spring.
Start
year: 1996
Location: Roadsides,
railroad rights of way, and nature preserves in and around Solem Township, MN
Overlaps
with: phenology in experimental plots, demography in the
remnants, gene flow in remnants, reproductive fitness in remnants
Data/materials
collected:We identify each plant with a numbered tag affixed to
the base and give each head a colored twist tie, so that each head has a unique
tag/twist-tie combination, or “head ID”, under which we store all phenology
data.We monitor the flowering status of all flowering plants in
the remnants, visiting at least once every three days (usually every two days)
until all heads were done flowering to obtain start and end dates of flowering.
We managed the data in the R project ‘aiisummer2019′ and added the records to
the database of previous years’ remnant phenology records, which is located
here: https://echinaceaproject.org/datasets/remnant-phen/.
A flowering curve (created here using the R package mateable) summarizes the flowering phenology data that we collected in 2019, indicating the number of individuals flowering on a given day and the flowering period for all individuals over the course of the season.
We shot
GPS points at all of the plants we monitored. Soon, we will align the locations
of plants this year with previously recorded locations and given a unique
identifier (‘AKA’). We will link this year’s phenology and survey records via
the headID to AKA table.
We
harvested a random sample of 1/3 of the flowering heads from each remnants in
August and September, plus an X additional heads from populations that were highly
isolated, for a total of X harvested seedheads. These are currently stored at
the University of Minnesota. This winter, I will assess the relationship
between phenology and reproductive fitness by x-raying all of the seeds we collected.
In addition, I will determine the paternity (i.e., pollen source) for a sample
of seeds by matching the seed genotype to the potential pollen donors. Doing so
will shed light on how phenology influences pollen movement and gene flow
patterns.
You can
find more information about phenology in the remnants and links to previous
flog posts regarding this experiment at the background
page for the experiment.
Products: I presented a poster based on the
locations and flowering phenology of individuals from summer 2018 at the
International Pollinator Conference in Davis, CA this summer. The poster is
linked here: https://echinaceaproject.org/international-pollinator-conference/.
Small remnant Echinacea populations may suffer from
inbreeding depression. To assess whether gene flow (in the form of pollen) from
another population could “rescue” these populations from inbreeding depression,
we hand pollinated Echinacea from six different prairie remnants with pollen
from a large prairie remnant (Staffanson Prairie) and from a relatively small
population that we call “Northwest Landfill.” We also performed
within-population crosses as a control. Amy Dykstra planted achenes (seeds)
that resulted from these crosses in an experimental plot at Hegg Lake WMA.
Plants in the crossing plots were originally found as seedlings like this one
We sowed a total of 15,491 achenes in 2008. 449 of these
achenes germinated and emerged as seedlings. Each summer we census the
surviving plants and measure them. This summer we found 48 surviving plants.
None of these plants has flowered, but we think some of them are close! The
largest plant we measured had 4 leaves, the longest of which was 35 cm.
You can read more about the interpopulation crosses, as well as links to prior flog entries mentioning the experiment, on the background page for this experiment.
Start year: 2008
Location: Hegg
Lake WMA
Data collected: Plant
fitness measurements (plant status, number of rosettes, number of leaves, and
length of longest leaf), and notes about herbivory. Contact Amy Dykstra to
access this data.
Products: Dykstra, A. B. 2013. Seedling recruitment in fragmented populations of Echinacea angustifolia. Ph.D. Dissertation. University of Minnesota. PDF