|
|
In 2015, we continued to gather demographic information on Echinacea angustifolia in remnant prairie populations. We went to 32 remnant prairies ranging size from 1 flowering plant to 289 flowering plants. This year at our largest site, the Staffanson Prairie Preserve, we collected demographic information on 318 individual plants including 175 flowering plants. Across all the sites, we found 1561 flowering plants and visited a total of 1889 plants. For each plant that we visit, we record whether the plant is alive, whether the plant is flowering, and how many flowering heads it produces.
We currently have former team member Lydia English and current team member Will Reed working on organizing the previous 20 years of data using methods that Stuart, Jared, and Gretel developed last year. So far we have organized data from 2010-2015 (years we used GPS units) in 10 sites and data for all 1995-2015 for Staffanson Prairie Preserve.
Read previous posts about this experiment.

Start year: 1995
Location: more than 30 remnant prairies in and near Solem Township, Minnesota
Overlaps with: fire and flowering at SPP, flowering phenology in remnants
Products:
Wagenius, S. 2006. Scale dependence of reproductive failure in fragmented Echinacea populations. Ecology 87:931-941. PDF | Supplemental Material
Project “demap” – organized survey, demography, and phenology data.
In 2015, we continued to assess survival and growth of plants that were found as seedlings in the springs of 2007 to 2013. This year we went to 13 sites and 142 parental plants. We found a total of 162 former seedlings with 48 of those at the site East Elk Lake Road. At first, the task of reading the maps and triangulating plant positions was hard for the team. We were worried we weren’t going to finish by the end of the field season. But, once we got the hang of it, we sped through with ease.
Read other posts about this experiment.

Start year: 2007
Location: Remnant prairies in central Minnesota
Overlaps with: demographic census in remnants
Products: Check the flog for preliminary results and annual reports.
 Plenty of basal leaves but no flowering in Amy Dykstra’s local adaptation experimental plots in 2015 In 2008, Amy Dykstra began an experiment to study how adapted Echinacea populations are to their local environments. She collected achenes from three populations distributed across a wide section of Echinacea angustifolia’s range, from Western South Dakota to our study site in Western Minnesota. She established a plot near each collection site where she sowed achenes from all sites. Since then, Amy has assessed survival and fitness traits of the individuals in her plots annually.
Amy expected to see increasing proportions of achenes emerging as seedlings with decreasing distance between the seed source and the sowing site. Instead she found that seedling emergence increased by sowing site from western to central South Dakota to Minnesota, and that at each site achenes from Minnesota and western South Dakota had higher seedling emergence than those collected at the central South Dakota site. This result suggests that there isn’t evidence of local adaptation in Echinacea seedling establishment, but that source population matters, potentially due to maternal effects.
After six years of tracking survival to fitness, however, there is some indication that local adaptation may play a role in later life stages. No individuals have flowered yet in any of the plots, which is somewhat surprising for Echinacea angustifolia individuals which are typically 5-7 years old before flowering for the first time. Amy will continue this study in 2016, so stay tuned for updates!
Start year: 2008
Location: Grand River National Grassland (Western South Dakota), Samuel H. Ordway Prairie (Central South Dakota), Staffanson Prairie Preserve (West Central Minnesota), and Hegg Lake WMA (West Central Minnesota).
Overlaps with: Dykstra’s interpopulation crosses
Team members who have worked on this project: Amy Dykstra, flog posts written by Amy may provide additional detail about activities associated the the development and continuing progress on this project.
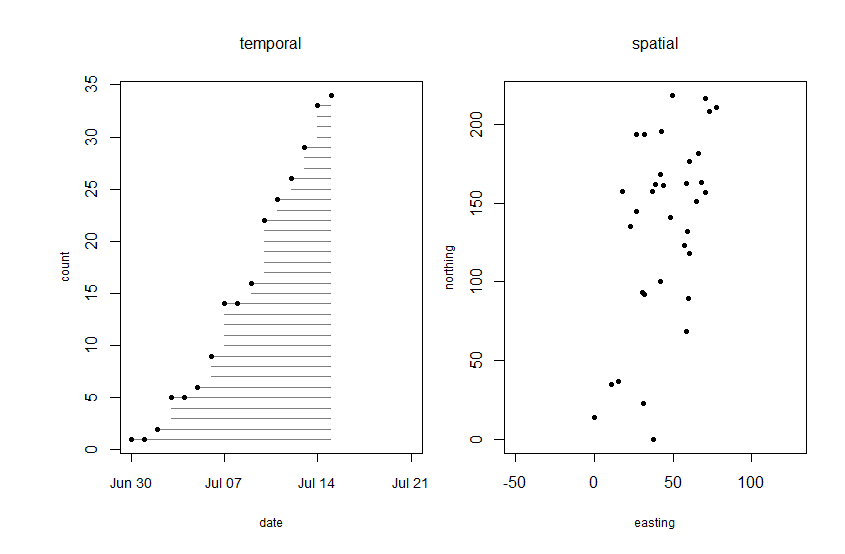
Echinacea pallida, a non-native Echinacea species that is compatible with E. angustifolia, was planted at a restoration at Hegg Lake WMA. One aspect of the potential that E. pallida has to interact with E. angustifolia individuals is the synchrony of their flowering timing, or phenology. To study this, we have kept track of the start and end dates of flowering for Echinacea pallida individuals in the Hegg restoration plot since 2011. In 2015, we identified 48 flowering plants with 139 heads. Flowering began on June 30th. Then, on July 15th, we chopped off all the Echinacea pallida heads. Before this date, 34 individuals flowered.
 Flowering schedule and map of Echinacea pallida at Hegg WMA. Note the abrupt end of flowering on July 15th. Click to enlarge! Read more about this experiment.
Start year: 2011
Site: Hegg Lake WMA restoration
Products: In Fall 2013, Aaron and Grace, externs from Carleton College, investigated hybridization potential by analyzing the phenology and seed set of Echinacea pallida and neighboring Echinacea angustifolia that Dayvis collected in summer 2013. They wrote a report of their study.
Overlaps with: Echinacea hybrids (exPt6, exPt7, exPt9), flowering phenology in remnants
Previous team members who have worked on this project include: Nicholas Goldsmith (2011), Shona Sanford-Long (2012), Dayvis Blasini (2013), and Cam Shorb (2014)
 Keke hard at work counting the q3 offspring Keke is a senior Environmental Studies major at Lake Forest College who has been working in the lab for the past semester. For her project, she focused on the maternal plants of qGen_3. In that experiment, we crossed individuals in p1 with pollen collected from plants at Staffanson and Landfill during the summer and planted the seeds from those crosses in the fall. When dissecting the heads, we only selected achenes that we knew had been crossed properly.
This left ray achenes and achenes that may have been contaminated with other pollen, plus any achenes that we missed! Although we didn’t want to plant inviable or contaminated achenes, knowing the fecundity of the maternal plants is an important part of estimating fitness, so we wanted to have accurate achene counts for each mom. This is where Keke comes in. She removed all of the “extra” achenes and counted them, along with the rest of the maternal achenes which had been scanned in the fall.
Keke also analyzed the effect of a new pollen management procedure that we followed for q3. This procedure involved collecting pollen in multiple vials and taking care to only remove a vial from the refrigerator for crossing once. This was in an effort to reduce exposure of pollen to repeated warming and cooling cycles, which we thought might have reduced its viability in q2. Keke assessed the percent of successful crosses in q2 versus q3 and found that the percent of successful crosses increased 5% with the new procedure. Cool!
You can read more about what Keke did this winter and spring in her report, which can be found here:
Keke’s Q3 Report
Thanks Keke and best of luck in all of your future endeavors!
In 2015, we continued an experiment that investigates hybridization between the native Echinacea angustifolia and the unintentionally planted non-native E. pallida. This year, out the the 758 plants planted in spring 2014, 521 were still alive which is a survival rate of 68.7%.
In the late summer of 2013, members of Team Echinacea collected heads from Echinacea angustifolia and Echinacea pallida from two nearby populations at Hegg Lake Wildlife Management Area. Unlike previous experiments, we performed no artificial crosses. This allows us to determine if hybridization is occurring naturally. In the winter of 2014, Lydia English germinated seeds from these heads. In the spring, Lydia and Stuart planted 758 seedlings at Hegg Lake WMA near experimental plot 7. We took fitness measurements, such as number of rosettes and leaf lengths, this year.
In addition to the experimental plot, we collected heads and tissue samples from 28 E. angustifolia that were near the restoration with E. pallida. We have not yet done any analysis on these plants but we are hoping to determine if hybridization in continuing.
Read more posts about this experiment here.
Echinacea Pallida at Hegg Lake Start year: 2014
Location: Hegg Lake Wildlife Management Area – Experimental plot 9
 A hybrid at Hegg Lake In 2015, we continued an experiment that quantifies fitness of Echinacea angustifolia x pallida hybrids and pure-strain plants. Out of the the original 522 plants, 323 were still alive in 2015, which is a survival rate of 62%. The mean leaf length of these plants was roughly 11.7 cm. Stuart planted the 522 seedlings at Hegg Lake WMA in spring 2013. The seedlings result from hand reciprocal crosses conducted by Shona Sanford-Long during the summer of 2012.
Read more posts about this experiment here.
Start year: 2013
Location: Hegg Lake Wildlife Management Area – Experimental plot 7
In 2015, we continued an experiment investigating fitness of Echinacea angustifolia x E. pallida hybrids. This year, out the the original 66 plants, 55 were still alive. That’s an impressive survival rate of 83% since they were planted in 2012. The mean leaf length of the plants was roughly 16 cm. In the summer of 2011, Nicholas Goldsmith and Gretel Kiefer performed reciprocal crosses between 5 plants of Echinacea pallida (non-native) found in a prairie restoration at the Hegg Lake Wildlife Management Area and 31 plants of the native Echinacea angustifolia from experimental plot 1 to determine the hybridization potential of these two species. In the summer of 2012, team members planted 66 seedlings.
Read more flog posts about this experiment here.
Echinacea Pallida on Hegg Lake Start year: 2012
Location: Experimental plot 6
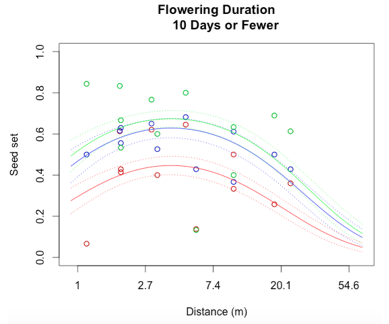
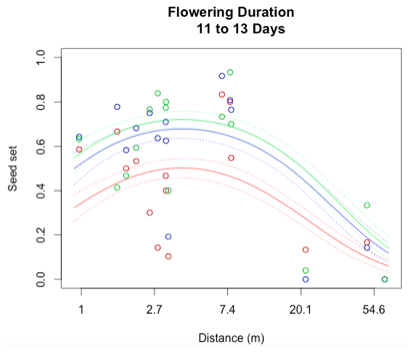
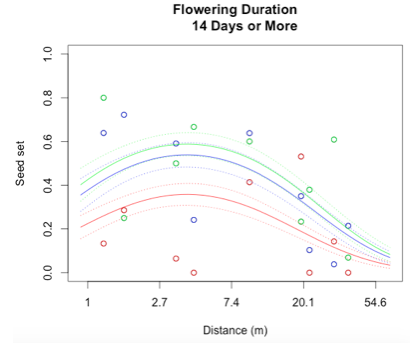
I successfully created a generalized linear model to describe reproductive success at Staffanson Prairie Preserve for the 2015 season. I started out with a model containing distance to the 3rd nearest neighbor, flowering start date, flowering duration, section of head from with the achene came, and all interactions between these main effects. In addition, I included both linear and quadratic terms for distance, start date, and duration. The purpose of including a quadratic term is to test for evidence of curvature in the relationship to seed set.
Through a process called backwards elimination, I removed all terms from the model that did not have a significant impact on reproductive success. The final model I came up with contained both linear and quadratic terms distance and start date, as well as the section of head. No interactions were found to be significant. In the plots below, the raw data are shown as points, and the predicted valued from the model are shown as curves. There are three plots of seed set as a function of distance, with the three plots representing low (10 days or fewer), mid (11-13 days), or high (14 days or more) flowering duration. The colors correspond to section of head, with red being top, blue is middle, and green is bottom.
  
While this is an observational study, so attributing causation would be inappropriate, it is helpful to think about why these relationships may exist. The general trend with increasing distance and spatial isolation is that seed set and reproductive success decrease. This is probably because isolated plants have fewer available mates or may be visited by fewer pollinators. However, the relationship is curved and plants in very densely populated areas also have diminished seed set. This may be caused by overcrowding and competition between plants for pollinators in areas of high population density.
Seed set has a similar peak at mid-duration of flowering. One explanation for this phenomenon is that plants that flower for a very short period of time co-flower with fewer potential mates, so a longer flowering time would maximize the number of compatible mates flowering at the same time. However, it is also known that extended flowering can be a sign that the head has not been receiving sufficient pollen, explaining the negative relationship between duration and seed set seen at higher durations.
In the future, this model will be compared to similar models from previous years during which Staffanson underwent controlled burns. Any differences seen between the models may be indicative of how the mating scene changes during the year after a burn.
 Reina, Pamela, and Mike with the photosynthesis machine used in Kittelson et al. (2015) In 2015, we continued to study the effects of inbreeding on Echinacea angustifolia fitness. This experiment was planted in 2006 where each plant was produced from one of three cross types: between maternal half siblings; between plants originating from the same remnant, but not sharing a maternal parent; and between individuals from different remnants. We continued to measure fitness and flowering phenology in these plants.
This year, of the original 1443 plants in INB2, 561 were still alive. Of the plants that were alive this year, 8.3% were flowering and 76.3% have never flowered – we’re still waiting! Among the plants that were flowering, mean head count was 1.53 heads, with a maximum of five heads.
Read more posts about this experiment here.
Start year: 2006
Location: Experimental plot 1
Overlaps with: Phenology and fitness in P1
Products: Fitness measurements were collected during our annual assessment of fitness in P1.
The following paper was published in summer 2015 based on fieldwork conducted in 2013.
Kittelson, P., S. Wagenius, R. Nielsen, S. Qazi, M. Howe, G. Kiefer, and R. G. Shaw. 2015. Leaf functional traits, herbivory, and genetic diversity in Echinacea: Implications for fragmented populations. Ecology 96:1877–1886. PDF
|
|